 |
ŚFiNiA
ŚFiNiA - Światopoglądowe, Filozoficzne, Naukowe i Artystyczne forum - bez cenzury, regulamin promuje racjonalną i rzeczową dyskusję i ułatwia ucinanie demagogii. Forum założone przez Wuja Zbója.
|
| Zobacz poprzedni temat :: Zobacz następny temat |
| Autor |
Wiadomość |
monitor/konto_usuniete
Usunięcie na własną prośbę
Dołączył: 07 Lut 2008
Posty: 656
Przeczytał: 0 tematów
Skąd: Konto usunięte na prośbę użytkownika. Patrz: przycisk WWW
|
 Wysłany: Śro 13:13, 25 Maj 2011 Temat postu: NIEREDUKOWALNA ZŁOŻONOŚĆ POD WZGLĘDEM FUNKCJI. Wysłany: Śro 13:13, 25 Maj 2011 Temat postu: NIEREDUKOWALNA ZŁOŻONOŚĆ POD WZGLĘDEM FUNKCJI. |
|
|
Na przykładzie dyskutowanej z Szymonem możliwości ewolucji pompy jonowej ATP-azy.
http://www.sfinia.fora.pl/genesis,36/nieredukowalna-zlozonosc-pseudonaukowa-koncepcja,5714.html#145932

Jak rozumieć pojęcie "NIEREDUKOWALNA ZŁOŻONOŚĆ POD WZGLĘDEM FUNKCJI" ?
Czy występowanie w organizmach białek "homologicznych" (w których występują pewne analogie [podobieństwa w ich dudowie]) dowodzą możliwości wyewoluowania układów nieredukowalnie złożonych ?
Choć Michael Behe wyjaśnił te kwestie już wielokrotnie, to jednak różni neodarwiniści (i ich zwolennicy ) często powtarzają jak mantry poglądy ,które zostały podważone (nawet wyrywając je z kontekstu opracowań zwolenników IP !) i w dalszym ciągu usiłują tymi motywami wspierać [b]swoją pseudo-naukową argumentację :
Wątek ten zakładam z tej przyczyny iż mimo wielokrotnego (setnego, tysięcznego...) tłumaczenia tej prostej zasady oponenci w dalszym ciągu powołują się na ten motyw. Żaden człowiek nie ma czasu , ani ochoty tłumaczyć w kółko te same rzeczy. Więc teraz wystarczy dać linka do tego tekstu i sprawa będzie załatwiona.
| Cytat: | http://www.nauka-a-religia.uz.zgora.pl/index.php?action=tekst&id=70
"(...)W końcu, zamiast pokazać, w jaki sposób ich teoria radzi sobie z
tym problemem, darwiniści starają się obejść problem nieredukowalnej
złożoności przy pomocy gierek słownych. Podczas debaty, sponsorowanej
przez American Museum of Natural History, która odbyła
się w kwietniu 2002 roku między zwolennikami i przeciwnikami teorii
inteligentnego projektu, Kenneth Miller rzeczywiście stwierdził (....)że
pułapka na myszy nie jest nieredukowalnie
złożona, gdyż jej podzbiory, a nawet każda osobna część, wciąż mogą
„funkcjonować” niezależnie od tego układu. Miller zauważył, że
drążek przytrzymujący z pułapki na myszy może służyć jako wykałaczka,
a więc nadal pełni „funkcję”, nie będąc częścią pułapki na
myszy.
Wszystkich części pułapki można użyć jako przycisku do
papieru – ciągnął dalej – więc każda z nich pełni jakieś „funkcje”. A
skoro każdy przedmiot, który posiada masę, może posłużyć jako przycisk
do papieru, to każda część czegokolwiek pełni swoją własną
funkcję. Czary mary, nie istnieje nic takiego jak nieredukowalna zło-
żoność.W taki oto prosty sposób wyjaśniono poważny problem dla
gradualizmu, który każde dziecko może dostrzec w systemach, takich
jak pułapka na myszy.Oczywiście, powyższe proste wyjaśnienie opiera się na ewidentnie
błędnym przekonaniu, wyraźnej dwuznaczności. Miller używa słowa
„funkcja” w dwóch różnych sensach.
Przypomnijmy sobie, że definicja nieredukowalnej złożoności mówi,
iż usunięcie jakiejś części
„powoduje, że system przestaje sprawnie funkcjonować”. Nie
wspominając o tym w swym wystąpieniu, Miller przenosi nacisk z
osobnej funkcji samego nienaruszonego systemu na kwestię, czy
możemy znaleźć inne zastosowanie (czy „funkcję”) dla niektórych
jego części.
Jeśli jednak usunie się jakąś część z przedstawionej przeze
mnie pułapki, to nie złapie ona już myszy. System faktycznie przestaje
sprawnie funkcjonować, a więc jest nieredukowalnie złożony – właśnie
tak jak napisałem. Co więcej, funkcje tak łatwo przypisywane
przez Millera częściom pułapki – przycisk do papieru, wykałaczka, łańcuszek
na klucze i tak dalej – mają niewiele, albo nic wspólnego z
funkcją całego układu – łapaniem myszy, a więc nie daje nam
to żadnej wskazówki dla wyjaśnienia, w jaki sposób funkcja systemu
mogła powstać stopniowo.
Miller nie wyjaśnił właściwie niczego. Pozostawiając problem pułapki na
myszy za sobą, Miller przeszedł
następnie do omówienia wici bakteryjnej – i ponownie odwołał się do
tego samego błędnego przekonania.
Jeżeli nie pozostało nic innego,
należy podziwiać tę zapierającą dech zuchwałość próby słownego obrócenia
kolejnego poważnego problemu darwinizmu na jego korzyść.
W ostatnich latach wykazano, że wić bakteryjna jest znacznie bardziej
skomplikowanym systemem niż dotąd sądzono. Działa ona nie tylko
jako urządzenie o napędzie obrotowym, ale w jej skład wchodzi także
wyszukany mechanizm transportujący białka z wewnątrz na zewnątrz
komórki, tworzące wierzchni fragment wici. Miller bez zmrużenia
oczu zapewnia, że wić nie jest nieredukowalnie złożona, gdyż pewnych
białek wici może brakować, a pozostała reszta – być może niezależnie
– może nadal transportować białka. (Białka podobne – ale nie
identyczne – do białek znajdowanych w wici występują w systemie
wydzielinowym typu III u niektórych bakterii). Miller ponownie popadł
w dwuznaczność, przenosząc nacisk z funkcji układu, który
działa jak maszyna o napędzie obrotowym, na zdolność podzbioru
tego systemu do transportowania białek przez membranę. Jednak, jak
argumentowałem, usunięcie części wici całkowicie odbiera temu układowi
zdolność do funkcjonowania jak maszyna o napędzie obrotowym.
Dlatego, niezgodnie z twierdzeniami Millera, wić rzeczywiście
jest nieredukowalnie złożona. Co więcej, funkcja transportowania
białek ma bezpośrednio tyle wspólnego z funkcją napędzania obrotowego,
ile wykałaczka z pułapką na myszy. Tak więc odkrycie dodatkowej
funkcji transportowania białek nie mówi nam niczego o tym,
jak procesy darwinowskie mogły złożyć maszynę o napędzie obrotowym.".
"(.....)Rozważmy hipotetyczny układ, w którym białka homologiczne do
wszystkich części nieredukowalnie złożonego mechanizmu molekularnego
z początku pełniły inne indywidualne funkcje w komórce. Czy
nieredukowalny system mógł w takim przypadku zostać złożony z
pojedynczych składników, które pierwotnie funkcjonowały osobno –
jak proponują niektórzy darwiniści? Niestety, jak pisałem w Darwin’s
Black Box, 13 zarysowany powyżej obraz znacznie upraszcza ten
problem (...)części układu molekularnego muszą automatycznie odnaleźć
siebie nawzajem w komórce. Nie może ich ułożyć pewien inteligentny
czynnik(...) Aby odnaleźć się wzajemnie w komórce, oddziałujące ze sobą części muszą
mieć powierzchnie ukształtowane tak, żeby bardzo dobrze do siebie
pasować (....) Pierwotnie jednak funkcjonujące
z osobna składniki nie miałyby komplementarnych powierzchni.
Wszystkie oddziałujące ze sobą powierzchnie wszystkich
składników musiałyby więc zostać dopasowane do siebie, zanim zaczęłyby
działać razem. Dopiero wtedy mogłaby pojawić się nowa
funkcja złożonego systemu. Dlatego mocno podkreślam, że problem
nieredukowalności nie znika, nawet jeśli pojedyncze białka homologiczne
do składników układu oddzielnie i pierwotnie pełniły swoje własne funkcje (......)
Nie należy też pochopnie wnioskować o redukowalności systemów
posiadających dodatkowe, czyli redundantne składniki, mogą one
bowiem posiadać nieredukowalnie złożony rdzeń. Na przykład, samochód
z czterema świecami zapłonowymi może jeździć z trzema lub
dwiema świecami, lecz z pewnością nie pojedzie bez żadnej. Pułapki
na szczury mają często dwie sprężyny w celu zwiększenia ich siły.
Taka pułapka może działać po usunięciu jednej sprężyny, ale nie
będzie działała, gdy usunie się dwie (.......) |
UZASADNIENIE NA PRZYKŁADZIE MATERIALNYM , TO ZNACZY ROZWAŻANEJ MOŻLIWOŚCI EWOLUCJI POMPY JONOWEJ ATP-azy (który to enzym jest ostatnim elementem dłuższego kompleksu enzymatycznego).


Neodarwiniści argumentują:
| Cytat: | " Cała siła argumentu Behe’ego opiera się na twierdzeniu, że
w biochemicznej maszynerii żywej komórki odkrył on jakąś nową własność (nieredukowalną złożoność), która umożliwia odrzucenie, nawet w zasadzie, jakiejkolwiek możliwości, że mogłaby ją wytworzyć ewolucja. Następne pytanie, jakie powinniśmy
zadać, jest proste – czy Behe ma rację?W roku 1998 Siegfried Musser i Sunney Chan opisali ewolucyjny rozwój pompy
protonowej oksydazy cytochromu c – złożonego, wieloczęściowego mechanizmu
molekularnego, który odgrywa kluczową rolę w przemianie energii w komórce.
W komórkach ludzkich pompa ta składa się z sześciu białek, z których każde jest konieczne
dla właściwego jej funkcjonowania. Wydaje się ona doskonałym przykładem
nieredukowalnej złożoności. Zabierz jedną część, a pompa nie będzie już pracować.
A mimo to autorzy mogli stworzyć, w imponujących szczegółach, „drzewo ewolucyjne,
skonstruowane dzięki wyobrażeniu, że złożoność i wydajność oddechowa zwiększały
się podczas procesu ewolucyjnego”. 8
Jak to możliwe? Gdybyście uwierzyli w zapewnienie Michaela Behe’ego, że mechanizmy
biochemiczne są nieredukowalnie złożone, nie chcielibyście tego sprawdzać
– i to jest prawdziwe naukowe niebezpieczeństwo jego idei. Musser i Chan sprawdzili
to i zobaczyli, że dwa z sześciu białek w pompie protonowej są całkiem podobne do
enzymu bakteryjnego, znanego jako kompleks cytochromów bo3. Czy może to oznaczać,
że część pompy protonowej wyewoluowała z działającego kompleksu cytochromów
bo3? Z pewnością – tak.
Ancestralny, dwuczęściowy kompleks cytochromów bo3 mógłby być w pełni funkcjonalny,
choć w innym układzie, lecz ów układ w rzeczywistości pozwalałby doborowi
naturalnemu na faworyzowanie jego ewolucji. Skąd pewność, że ta „połowa”
pompy w ogóle byłaby użyteczna? Mamy ją dzięki odniesieniu się do współcześnie
żyjących organizmów, które mają pełną, działającą wersję kompleksu cytochromów
bo3. Czy możemy użyć tego samego argumentu dla pozostałej części tej pompy? Cóż,
okazuje się, że każda z głównych części pompy jest blisko związana z działającymi
kompleksami proteinowymi, znajdującymi się w mikroorganizmach.
Jak wskazali Musser i Chan, ewolucja kompletuje złożone mechanizmy biochemiczne z mniejszych
działających mechanizmów biochemicznych, które zostały przystosowane do pełnienia
nowych funkcji. Wiele części złożonych mechanizmów biochemicznych składa się
z mniejszych działających mechanizmów, rozwiniętych przez dobór naturalny (....)" ] |
Do wyjasnienia mozliwości ewolucji pompy jonowej nie wystarczy znalezienie (ewentualnych) homologów [analogów]. Ona się składa z dwóch modułow (Fo i F1)
1) oraz z precyzyjnego aparatu (coś jak stator w silniku elektrycznym) do selektywnego dawkowania jonów umozliwiajacych rotorowi ruch obrotowy ,oraz powrót jonów do wnętrza komórki.

Konformacja w tym przypadku musi być idealna ,a kanały jonowe idealnie wymierzone, co widać na przekroju w zamieszczonym modelu !
2) Enzymy z kompleksu F1 muszą mieć odpowiednią konformację
Zeby się odpowiednio dopasować do siebie samych
3) Zeby wał łaczacy kompleks Fo z F1 mógł wchodząc jednym końcem pomedzy te białka i obracając się mógł powodować zmianę ich konformacji (kształtu) , co dopiero umozliwia efektowną syntezę ATP z ADP (pod względem FUNKCJI jaką pełni pompa jonowa ATP-azy) !

Najprostszy kompleks do rotacyjnej syntezy ATP składa się z 8 podjednostek.
5 z tych podjednostek wchodzi w sklad katalitycznego kopleksu F1
(AlFa, Beta, Gamma, Delta i Eps.). Napędzana przez protony Podjednostka Fo
składa się z 3 rodzajów podjednostek (a, b ,c).
Katalityczna część syntazy ATP (F 1) utworzona jest przez podjednostki Alfa 3 i Beta 3 tworząc heksamer.

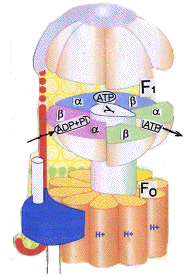
Podjednostki gamma i delta są związane od "góry" i "dołu" z heksamerem, podjednostka gamma dodatkowo łączy się z podjednostką b2 . Hydrofobowy segment transbłonowej podjednostki b2 jest w kontakcie z podjednosteką a. Przeniesienie protonu odbywa się na styku podjednostki A i C.

Na zmianach w konformacji róznych enzymów opiera się praktycznie całe funkcjonowanie zywej komórki.
Zmiany w konformacji białek wywoływane są w przerózny sposób. W przypadku pompy jonowej ATP-azy (naprzemienne )zminany konformacji w module F1 wywoływane sa mechanizcznie poprzez przedłużone oddzialywanie rotora napedzanego strumieniem jonów. To przedłużenie to wspomniany wał ,który także musi mieć odpowiednią konformację (kształt).
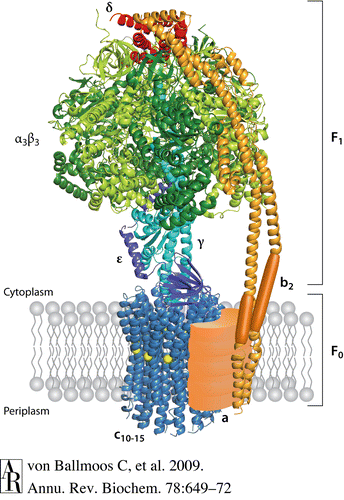
Zeby można było (ewentualnie ) uzyskać odpowiednie domeny (miejsca w białkach , które umozliwiają im wzajemne, subteone dopasowania) należy zakładać dlugą ewolucję. W przypadku kompleksu rotacyjnego ATP-azy takie subtelne dopasowania, pod względem FUNKCJI tego molekularnego urządzenia, musiały być DOKLADNE już od samego początku istnienia tego kompleksu ,a więc dobór naturalny musiałby (ewentualne) 'białka homologiczne' niejako 'proroczo' dopasowywać do przyszłej fukcji ,a następnie stopniowo dołaczać do coraz bardziej złożonego układu, jakim jest kompleks rotacyjny ATP-azy. Co jest NIEMOZLIWE do przyjecia , ponieważ ten kompleks POD WZGLĘDEM funkcji OD RAZU musi być kompleksem KOMPLETNYM.
Nawet (NAWETT!!!) jeżeli okazałoby się iż w jednej bakterii występują WSZYSTKIE "homologii" , to znaczy analogii dla kompleksu ATP-azy, które by pełniły inne funcje, a nawet te z komleksu F1 , które syntetyzowałyby ATP z ADP, lecz nie w wyniku mechanicznej zmiany konformacji, jak jak to się dzieje w kompleksie rotacyjnym F1 ATP-azy, ale posiadając jedynie domenę umożliwiajacą syntezę ATP z jakiegoś substratu (podobne występują), to i tak takie białka zanim nabyłyby odpowienich domen ; KONIECZNYCH MOŻLIWOŚCI ABY SIĘ DOPASIWAĆ do siebie na wzajem, do sibie i do wału , ktory obracając się wywołuje zmiany ich kształtu, byłyby dla fukcji kompleksu rotacyjnego ATP-azy BEŻUZYTECZNE.
Powraca więc pytanie postawione przez Webera: w jaki sposób układy niekompletne (w tym przypadku pompa jonowa) mogły zdobywać przewagę selekcyjną ? Te "bardzo podobne" białka cytochromowe z komleksu bo 3 są podobne , ale nie identyczne, do tych (subtelnie dopasowanych do pełnienia odpowiedniej funkcji), które wchodzą w skład kopleksu rotacyjnego ATP-azy.
Jak by to się wogóle mogło stać , żeby te tak odmienne, pierwotne białka (prekursorzy) mogły zaczać pełnić tak odmienną funckcję ? Scenariusz neodarwinowski (i jedynie logiczny na tej kanwie do przyjęcia) może być tylko jeden.
1) Więc najpierw musiałaby zajść duplikacje ich genów. 2) Następnie produkcja IDENTYCZNYCH, w stosunku do formy wyjściowej, białek , co mogłoby (fantazjując) nastąpić
poprzez dokopowanie ich pod jakiś regulacyjny mechanizm genetyczny , który by powodował ich transkrypcję. 4) Następnie musiałaby się zacząć STOPNIOWA dywergencja tych genów w kierunku ewolucji dopasuwującej te geny pod kątem funkcji pełnionej w kompleksie rotacyjnym ATP-azy.
Nawet jeżeli zdrowy rozsądek pozwoli komuś przyjąć , iż te dopasowane do siebie domeny wyewoluowaly niezależnie w białkach, które wcześniej (już po duplikacjach) pełniły odmienne funkcje niż ta funkcja , którą pełnią obecnie w kompleksie rotacyjnym ATP-azy. To i tak musiałoby zbiec się to wszystko w jednym czasie (łącznie z wyewoluowaniem odpowiednich genetycznych czynnków regulujących transkrypcję i translacje genów kodujących te białka [odpowiednia regulacja w transkrypcji ilości kopii danego białka, kolejności transkrypcji itd.]). W przeciwnym razie pompa jonowa rotacyjnego kompleksu ATP-azy nie mogłaby zostać skompletowana ! Więc ludzie odrzucajacy IP i nieredukowalną złozoność są ZMUSZENI (po prostu nie pozostaje im zadna inna opcja dowyboru) uznać iż pompa jonowa kompleksu rotacyjnego ATP-azy powstała za jednym zamachem.
Bez uczestnictwa w takim kompleksie od samego początku dobór nie mógłby dopasowywać praktycznie zadnego z elementów pompy rotacyjnej ATP-azy pod wzgędem pełnionej przez to molekularne urzadzenie funkcji ! W JAKI SPOSÓB mógłby np. ukształtować odpowiedni KSZTAŁT wała , który napedza całą syntezę ATP, poprzez przenoszenie ruchu obtotowego do kompleksu F1 z kompleksu Fo, zmieniajac konformacje enzymów ktore tworzą kompleks F1?
W jaki sposób białka tworzace kompleks Fo mogły się do siebie dopasować i zdobyć wymierzone domeny , które umożliwiają wiązanie jonów (w odpowiedniej ilości !) , zmianę ich konformacji i ruch obrotowy całego rotora ?
W jaki sposób 'staor' zdobył odpowiednią konformację , żeby odpowiednio (SUPER-DOKŁADNIE!) pasować jednym końcem (od strony błony) do rotora, co umozliwia mu odpowiednie dawkowanie jonów (i ich odprowadzanie), a z drugiego końca odpowiednie przyleganie do zmieniających cyklicznie konformację enzymów z kompleksu F1 ?
W jaki sposób dobór naturalny mógł widzieć te geny i je do tej funkcji dopasowywać, skoro one jeszcze nie uczesniczyły w syntezie ATP; pełniąc odpowiednie funkcje w tym kompleksie rotacyjnym ATPazy ? I tutaj własnie pojawia się zasadnicze pytanie: "czy dobór naturalny działał proroczo ?"
Czy pompa jonowa tworząca komleks ATP-syntezy mogła działać jako komplekst niekompletny i wykonywać jakąkolwiek użyteczną funkcję w organizmie komórki ?
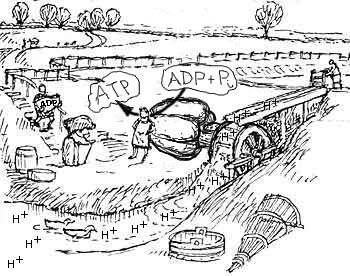

Czy można naciąć drzewa ,kiedy z elektrycznego silnika usunie się magnez (staror) lub rotor ? Czy kompleks silnik elektryczny-piła tarczowa połączone za pośrednictwem pasa tnansmisyjnego (odpowiednik wału w kompleksie ATP-azy) będzie dalej działać po usunięciu tego pasa ?
Pewne eksperymenty wykazały, że jeśli podjednostka gamma jest mechanicznie zmuszona do obrotu, synteza ATP zachodzi nawet bez transportu protonowego w module F O.
Ewolucjoniści z tym doświadczeniem wiązali duże nadzieje, to znaczy przypuszczali iż uda się udowodnić redukowalny charakter tej zadziwiajacej struktury.
Zakładano , ze taki (w odłaczeniu od rotora [podjednostki 'c']) ruch obrotowy samej podjednostki gamma może się odbywać w warunkach in vivo.
Jednak nie ma żadnych eksperymentalnych dowodów na obrotowy mechanizm takiego (wybrakowanego) typu w nienaruszonym kompleksie rotacyjnej ATP-azy (w warunkach fizjologicznych). NIE MA żadnych dowodów na to , że bez sztucznej "pomocy", w warukkach naturalnych, kompleks rotacyjny ATP-azy może prawidłowo funkcjonować inaczej niż jako układ kompletny napędzany przez przepływ jonów !
Jak widać , kiedy podda się gruntownej analizie większość neodarwinowskich bajeczek , które Ty traktujesz jako poważne hipotezy naukowe , to tak na prawdę nic z nich godnego uwagi nie pozostaje (biorąc pod uwagę "tysiace [fikcyjnych] artykułow naukowych", które rzomo obajśniają ewolucję układów nieredukowalne złozonych i na ktore powołują się namiętnie ewolucjoniści). No coż widzę , że do opracowania dotyczącego możliwości/niemożliwości ewolucji wici bakteryjnej jestem zmuszony dopisać rozdział poświęcony niemożliwości STOPNIOWEJ ewolucji prostszych , molekularnych, urzadzeń sekrecyjnych.
TO WIDAĆ NAWET NA OKO- np. na filmiku ,który możesz sciągnąć tutaj :
[link widoczny dla zalogowanych]
Tutaj dodatkowe informacje:
[link widoczny dla zalogowanych]
[link widoczny dla zalogowanych]
[link widoczny dla zalogowanych]
[link widoczny dla zalogowanych]
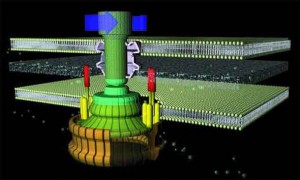
UZUPEŁNIENIE-HIPOTEZA MODULARNA W EWOLUCJI POMPY JONOWEJ KOPLEKSU ROTACYJNEGO ATP-azy.-
Syntaza ATP



Owiele bardziej skomplikowana Wic bakteryja dla porownania:



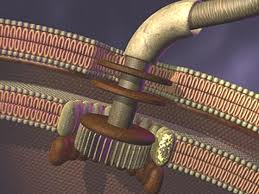
Sam element napędowy witki jest bardzo skomplikowany:

A żeby nie być gołosłownym ,to zaprezetuję "hipotezę" ,która postuluje ewolucyną genezę rotacyjnego kompleksu do sytnezy ATP. Mozna tą bajeczke przyrównać do twierdzenia : podzespoły (moduły) tego kompleksu powstały z wczesniejszych modułow ,ktore pełniły inną funckje i same były nieredukowalnie złozone.. Najbardziej zapawne jest w tym oswiadczeniu twierdzenie , ze odnosnie "modułu Fo to napedzajacy go przepływ jonów jest bardzo podobny do modułu napedzajacego wić bakterujną". Tak tylko ,co to wyjasnia ? Nic. Po pierwesze dlatego ,ze 'analogiczny moduł lokomocyjny' u bakterii jest o wiele bardziej skomplikowany, po drugie przecież ten 'analogiczny moduł' u e.coli ,wchodzacy w skład wici, miał przeciez powstać z prostszego modułu (takiego ,jaki występuje w kompleksach białkowych służacych do syntezy ATP['Fo']). Dobrze więc , ze autorzy wspomnieli tylko o 'podobieństwach' w napędzie jonowym.
No najwazniejsze pytanie. Jaką funkcję mógł pełnić sam moduł 'Fo' zanim połaczył się z resztą kompleksu do syntezy ATP ? On i dzisiaj sam moze się kręcic w kółko ,ale w jakim celu taki niekopletny uklad mógł wyewoluować , jaką dawać przewagę selekcyjną ,zeby dobór naturalny mógł wogóle tolerować tak kosztowną energetycznie (i bezuzyteczną) cechę ? No i w koncu--w jaki sposób moduł 'Fo' mógł wyewoluować skoro sam sklada się z kilku zintegrowanych elementów, które w ewolucji musiały pojawiać się stopniowo w historii 'ewolucji tej struktury' ?
Moduł Fo (podjednostek α i β ) po usunięciu modułu F1-w dalszym ciagu wykonuje ruchy obrotowe ,ale traci swoją funkcję.
(Sam jest jednak nieredukowalnie złozony).

Oto co sami ewolucjonisci maja na ten temat do powiedzenia:
[link widoczny dla zalogowanych]
| Cytat: | "Uważa się, że ewolucja syntazy ATP zachodziła modułowo. Obie domeny wraz ze swoimi wartościowościami połączyły się zyskując nową funkcję. Domena F1 wykazuje znaczne podobieństwo do heksamerycznej helikazy DNA, a domena Fo jest podobna do kompleksów tworzących motor molekularny napędzający wici komórek.
Heksamer α3β3 tworzący domenę F1 podobnie jak helikaza DNA składa się z obracającego się pierścienia z otworem po środku. U obu enzymów obroty pierścienia umożliwiają pełnienie funkcji. Helikaza DNA porusza się po helisie DNA przy jednoczesnej hydrolizie nukleotydów. Domena F1 wykorzystuje zmiany konformacyjne podczas obrotu podjednostki γ do przeprowadzenia reakcji enzymatycznej.
Jony H+ przepływające przez domenę Fo w bardzo podobny sposób jak przy napędzaniu motoru molekularnego poruszającego wicią. Ich wspólną cechą jest pierścień składający się z wielu alfa-helikalnych białek, które obracają się w stosunku do innych białek zużywając jednocześnie gradient protonowy jako źródło energii. Jest to jednakże dość wątły związek, ogólna struktura motorów molekularnych jest zdecydowanie większa, zawierają 30 polipeptydów, w porównaniu do 10, 11, lub 14 znanych polipeptydów domeny 'Fo'.
Modułowa teoria pochodzenia syntazy ATP sugeruje, że dwie domeny o niezależnych funkcjach, helikaza DNA posiadająca właściwości ATPazy i motor molekularny napędzany siłą protonomotoryczną, mogły się połączyć i doprowadzić do odwrócenia właściwości ATPazy powstałej z helikazy DNA. Dalszy rozwój prowadziłby do powstania kompleksy syntazy ATP znanej dzisiaj. Alternatywnie kompleks helikazy DNA i motoru molekularnego mógł wykazywać początkowo aktywność ATPazy, która przenosiła jony H+ zużywając ATP. Dalsza ewolucja kompleksy mogłaby doprowadzić do odwrócenia przeprowadzanej reakcji i powstania funkcji spełnianej przez syntazę ATP."
|
Koncepcja ewolucji modułu F 1 od struktury podobnej do helikazy DNA
też jest z resztą zwykłą bajeczką:

Coś takiego ('moduł F1' z kompleksu suntetyzującego ATP):

Musiałoby wyewoluować z czegoś takiego ,jak helikaza DNA, która pełni
inna funkcję i zawiera (poza podobieństwami) duzo róznic strukturalnych i
biochemicznych w porównaniu z 'modułem F 1.

Innymi słowy-1 enzym (Moduł F 1) jest przystosowany do syntezy ATP w takim kompleksie, w jakim
uczestniczy w tym procesie.
2 enzym( Helikaza DNA) jest przystosowany do rozplatania DNA podczas jego replikacji i napraw. Mimo podobieństwo isnieje pomiedzy tymi enzywami tyle roznic ,co do biochemi i struktury domen, jak i konformacji ,ze nie sposob szukać tutaj wyjasnia w homologii (czy chocby analogii).


(.....)"
pozdrawiam;
Ostatnio zmieniony przez monitor/konto_usuniete dnia Śro 23:06, 25 Maj 2011, w całości zmieniany 9 razy
|
|
| Powrót do góry |
|
 |
|
|
|
| Zobacz poprzedni temat :: Zobacz następny temat |
| Autor |
Wiadomość |
awaryjan
Dołączył: 05 Maj 2011
Posty: 53
Przeczytał: 0 tematów
Płeć: Mężczyzna
|
| Wysłany: Śro 21:38, 25 Maj 2011 Temat postu: |
|
|
Witam.
Mam dwa pytania związane z tematem nieredukowalnej złożoności, tak bardziej z ciekawości:
1) Behe z tego co pamiętam jako jeden z przykładów podawał proces krzepnięcia krwi. Wiadomo jednak, iż istnieje co najmniej kilka wersji tego procesu, np. delfinom nie jest do niczego potrzebny czynnik Hagemana, a krzepnięcie i tak zachodzi, są jeszcze prostsze przykłady tego procesu. Z tego wynika, że nasze krzepnięcie jednak jest nieco redukowalne. Jak to podciągnąć pod "nieredukowalność"? W którym miejscu "nieredukowalność" by się skończyła twoim zdaniem?
2) wspominasz o wici bakteryjnej. Z pewnością znasz model zaproponowany przez Matzkego. Jest jaki jest, ale moim zdaniem sporo popchnął do przodu w temacie ewolucji tego elementu. Gdzie dostrzegasz najważniejsze jego uchyby?
Chodzi mi tylko o zarys, krótkie podsumowanie tych zagadnień, bez odwoływania się do masy materiału (tam, gdzie się da). Z góry dzięki.
|
|
| Powrót do góry |
|
|
| Zobacz poprzedni temat :: Zobacz następny temat |
| Autor |
Wiadomość |
monitor/konto_usuniete
Usunięcie na własną prośbę
Dołączył: 07 Lut 2008
Posty: 656
Przeczytał: 0 tematów
Skąd: Konto usunięte na prośbę użytkownika. Patrz: przycisk WWW
|
| Wysłany: Śro 22:42, 25 Maj 2011 Temat postu: |
|
|
Również witam !
| Cytat: | Mam dwa pytania związane z tematem nieredukowalnej złożoności, tak bardziej z ciekawości:
1) Behe z tego co pamiętam jako jeden z przykładów podawał proces krzepnięcia krwi. Wiadomo jednak, iż istnieje co najmniej kilka wersji tego procesu, np. delfinom nie jest do niczego potrzebny czynnik Hagemana, a krzepnięcie i tak zachodzi, są jeszcze prostsze przykłady tego procesu. Z tego wynika, że nasze krzepnięcie jednak jest nieco redukowalne. Jak to podciągnąć pod "nieredukowalność"? W którym miejscu "nieredukowalność" by się skończyła twoim zdaniem ? |
Znam temat tylko pobieżnie. Szczerze mówiąc nie czytałem tej pracy Dollittla, choc była napisana dawno temu. Po prostu kaskada krzepnięcia krwi leży na granicy moich zainteresowań. Biochemia tego procesu jest mi znana pobieżnie, więc nie będę się wymądrzał. Michael Behe coś na pewno na ten temat napisał ,ja ostatnio (jakiś miesiąc temu ) czytałem krótką przegladówkę na jakiejś stronie przeciwników NZ. Powiem Ci jednak szczerze ,ze przeszła mi przez głowę myśl ,zeby się tym zająć, ale jak sobie pomyślę o złozoności tego procesu ,to zaczyna mi się kręcić w głowie :-).
Jutro poszukam coś w sieci chodząc swoimi sciezkami. Jeżeli Behe coś napisal na ten temat ,to podam Ci linka. Ze swojej strony (luzna uwaga) mogę powiedzieć ,że ryby bezszczękowe nie muszą mieć prostszej kaskady ,ponieważ może u nich ona przebiegać w inny sposób. Alternatywne szlaki tego procesu mogą być wyposazone w odmienne białka. Np. istnieje około 10 alternatywnych szlaków rozkładu glukozy, a jednak jakby to wszystko zredukować do kilku szlaków ,czy nawet jednego ,to i tak okazuje się on nieredukowalnie złozony.
Prostsze kaskady to nie nowość, bo np. bezkręgowce (skorupiaki) mają zamiast krwi tzw hemolimfę i przy takim składzie tej "prymitywnej krwi" i otwartym układzie krwionośnym wystarcza im protsza kaskada. Jednak nie można jej traktować jako prekursora tej ,która występuje u kręgowców.
Behe zajmuje się biochemią białek ,więc na pewno wie więcej na ten temat. Napisz do niego jak nic nie znajdę.
EDIT :
Nie trzeba było dlugo szukać ,no i ja sam się nie pomyliłem stawiając swoją ogólną hipotezę. Zacutuję tutaj fragment ,a Ty idz tym tropem, a dowiesz się więcej (linków w tekście):
[link widoczny dla zalogowanych](Talk.Origins)
| Cytat: | " Behe's Response to this Objection
In fact, this objection to Behe’s argument for the irreducible complexity of the blood-clotting cascade is entirely beside the point. In the Kitzmiller v. Dover case, Behe, in his testimony, dealt explicitly with this ‘rebuttal’ as expressed by Dr. Ken Miller. Here is the relevant portion of Behe’s testimony, running from page 25, line 9 to page 30, line 5 (A = Behe):
* Q. -- and Doolittle and Davidson, et al, to argue against the irreducible complexity of the blood clotting system. Do you agree with his assessment of those studies?
* A. No, I do not.
* Q. And you have some diagrams to explain this further, sir?
* A. Yes, I do. This is a slide from Professor Miller's presentation showing work from Jiang and Doolittle. And he also shows a diagram of the blood clotting cascade. And notice again, it's a branched pathway with the intrinsic pathway and the extrinsic pathway. And Professor Miller makes the point that in DNA sequencing studies of something called a puffer fish, where the entire DNA of its genome was sequenced, and scientists looked for genes that might code for the first couple components of the intrinsic pathway, they were not found.
And so Professor Miller demonstrated that by -- if you could push to start the animation -- Professor Miller demonstrated that by having those three components blanked out in white. Nonetheless, puffer fish have a functioning clotting system. And so Professor Miller argued that this is evidence against irreducible complexity.
But I disagree. And the reason I disagree is that I made some careful distinctions in Darwin's Black Box. I was very careful to specify exactly what I was talking about, and Professor Miller was not as careful in interpreting it. In Darwin's Black Box, in the chapter on blood clotting cascade, I write that, a different difference is that the control pathway for blood clotting splits in two. Potentially then, there are two possible ways to trigger clotting.
The relative importance of the two pathways in living organisms is still rather murky. Many experiments on blood clotting are hard to do. And I go on to explain why they must be murky. And then I continue on the next slide. Because of that uncertainty, I said, let's, leaving aside the system before the fork in the pathway, where some details are less well-known, the blood clotting system fits the definition of irreducible complexity. And I noted that the components of the system beyond the fork in the pathway are fibrinogen, prothrombin, Stuart factor, and proaccelerin. So I was focusing on a particular part of the pathway, as I tried to make clear in Darwin's Black Box. If we could go to the next slide. Those components that I was focusing on are down here at the lower parts of the pathway. And I also circled here, for illustration, the extrinsic pathway. It turns out that the pathway can be activated by either one of two directions. And so I concentrated on the parts that were close to the common point after the fork.
So if you could, I think, advance one slide. If you concentrate on those components, a number of those components are ones which have been experimentally knocked out such as fibrinogen, prothrombin, and tissue factor. And if we go to the next slide, I have red arrows pointing to those components. And you see that they all fall in the area of the blood clotting cascade that I was specifically restricting my arguments to. And if you knock out those components, in fact, the blood clotting cascade is broken. So my discussion of irreducible complexity was, I tried to be precise, and my argument, my argument is experimentally supported.
* Q. Now just by way of analogy to maybe help explain further. Would this be similar to, for example, a light having two switches, and the blood clotting system that you focus on would be the light, and these extrinsic and intrinsic pathways would be two separate switches to turn on the system?
* A. That's right. You might have two switches. If one switch was broke, you could still use the other one. So, yes, that's a good analogy.
* Q. So Dr. Miller is focusing on the light switch, and you were focusing on the light?
* A. Pretty much, yes.
* Q. I believe we have another slide that Dr. Miller used, I guess, to support his claim, which you have some difficulties with, is that correct?
* A. Yes, that's right. Professor Miller showed these two figures from Davidson, et al, and from Jiang, et al, Jiang and Doolittle, and said that the suggestions can be tested by detailed analysis of the clotting pathway components. But what I want to point out is that whenever you see branching diagrams like this, especially that have little names that you can't recognize on them, one is talking about sequence comparisons, protein sequence comparisons, or DNA nucleotide sequence comparisons. As I indicated in my testimony yesterday, such sequence comparisons simply don't speak to the question of whether random mutation and natural selection can build a system. For example, as I said yesterday, the sequences of the proteins in the type III secretory system and the bacterial flagellum are all well-known, but people still can't figure out how such a thing could have been put together. The sequences of many components of the blood clotting cascade have been available for a while and were available to Russell Doolittle when he wrote his essay in the Boston Review. And they were still unhelpful in trying to figure out how Darwinian pathways could put together a complex system. And as we cited yesterday, in Professor Padian's expert statement, he indicates that molecular sequence data simply can't tell what an ancestral state was. He thinks fossil evidence is required. So my general point is that, while such data is interesting, and while such data to a non-expert in the field might look like it may explain something, if it's asserted to explain something, nonetheless, such data is irrelevant to the question of whether the Darwinian mechanism of random mutation and natural selection can explain complex systems.
* Q. So is it your opinion then, the blood clotting cascade is irreducibly complex?
* A. Yes, it is. " |
Z mojej strony kończę dyskusje na temat kaskady krzepnięcia. Zajmuję się obecnie układami sekrecyjnymi i jeszcze to dość długo potrwa. Innymi słowy nie mam czasu na wchodzenie głębiej w innego rodzaju zagadnienia. Być może kiedyś powrócę do tego tematu (może Ty postarasz się mnie zainteresować jakimś zgrabnym elaboratem prezentującym oba stanowiska i wyłuszczajacym Twoje wnioski ?).
| Cytat: | | 2) wspominasz o wici bakteryjnej. Z pewnością znasz model zaproponowany przez Matzkego. Jest jaki jest, ale moim zdaniem sporo popchnął do przodu w temacie ewolucji tego elementu. Gdzie dostrzegasz najważniejsze jego uchyby? |
Tym modelem obecnie się zajmuję. Więcej o moich planach przeczytasz w watku załozonym przez Szymona:
http://www.sfinia.fora.pl/genesis,36/nieredukowalna-zlozonosc-pseudonaukowa-koncepcja,5714.html
Przedostatni post. Widzisz nie mogę z tym ruszyć całą parą choć matrialy zgromadzone ,wiekszość szkiców, bo cały czas coś .Teraz Szymon wezwał mnie do tablicy w związku z pompą protonową ATP-azy, więc wertując internet pół nocy i cały dzeń napisałem powyższy post. Ale OK, teraz jak widać neodarwinizm ma problem nie tylko z ewolucją samej witki ,ale również z ewolucją prostszych "prekursorów" od których owa ewolucja miała wystartować.
| Cytat: | | Chodzi mi tylko o zarys, krótkie podsumowanie tych zagadnień, bez odwoływania się do masy materiału (tam, gdzie się da). Z góry dzięki. |
Powiem Ci szczerze ,ze kiedy zaczynam pisać na ten temat jakiś post ,to zawsze zakladam ,ze zajmie mi to ze trzy godziny. Pozniej się okazuje ,ze jak kończę (albo czasami jestem w połowie), to zaczyna switać. Pomyśl sam : zeby opisać niemożliwość ewolucji układu (pompy jonowej ATP-azy) nalezy przewertować w zdłuz i szez internet i poświęcić na to prawie dwie doby. Nalezy sie zapoznac z tym ,co mają do powiedzenia ewolucjoniści,skonfrontować to z tym ,co mowi doswiadcznie i dopiero mozna uczciwie to wszystko podsumować. A przecież ten komplekst składa się z 9 elementów. Jakże więc mogę w kilku słowach opisać i obalić model Matzkego, choć wiekszość tego modelu zajmują opisy "homologów" oraz ilustracje pogladowe ?
Witka bakteryjna jest o wiele bardzioej zlozonym organem i cały czas nas zaskakuje. Pojawiają się na temat jej badań coraz nowsze i zdumiewajace doniesienia [link widoczny dla zalogowanych] . Ostatnio posciągałem swietne artykuły o sposobach montażu tego narządu (nokautowaniu genów), a zaraz pózniej trafiłem na angielską stronę japońskich uczonych, którzy zajmują się badaniem wici i trafiłem na jeszcze ciekawsze doniesienia. Badz cierpliwy, chce to zrobić rzetelnie. Jezeli mój materiał kogoś tam nie przekona do IP, to przynajmniej zdobędzie jakies wartosciowe informacje na temat tego ogranu.
Tak szczerze pisząc ,to model Matzkego jest już mocno przestarzały. Ale z braku w to miejsce czegoś lepszego (stara praktyka zwolenników SAModziejstwa) jest on dalej promowany. Nie mam z czym dyskutować ,więc i ja z braku laku musze dyskutować z tym modelem:-).
pozdrawiam;
Ostatnio zmieniony przez monitor/konto_usuniete dnia Czw 10:10, 26 Maj 2011, w całości zmieniany 5 razy
|
|
| Powrót do góry |
|
|
|
|
Nie możesz pisać nowych tematów
Nie możesz odpowiadać w tematach
Nie możesz zmieniać swoich postów
Nie możesz usuwać swoich postów
Nie możesz głosować w ankietach
|
fora.pl - załóż własne forum dyskusyjne za darmo
Powered by phpBB © 2001, 2005 phpBB Group
|

